Module 2. Classification of dairy microorganisms
Lesson 10
CHARACTERISTICS OF DAIRY ASSOCIATED FUNGI AND BACTERIOPHAGES
10.1 Introduction
Yeast and moulds play an important role in dairy, especially as spoilage micro-organisms. Some of the yeast and moulds are also used as starter cultures in the preparation of different fermented dairy products. Yeasts are unicellular in nature and grow by budding. Moulds are the only multicellular microorganism,in nature.
The fungi and bacteriohages that are signicant from dairy point of view are discussed here in this lesson. Fungi are classified as:
· Division - Mycota
· Sub Division - Mycotina
· Class - Mycetes
· Sub class - Mycetidae
· Order - ales
· Family - aceae
10.2 Moulds
Moulds (or molds) are fungi that grow in form of multicellular filaments called hyphae. A connected network of these tubular branching hyphae has multiple, genetically identical nuclei and is considered a single micro-organism, referred to as colony. In contrast, fungi that grow as single cells are called yeasts. Moulds do not form a specific taxonomic or phylogenetic grouping, but can be found in the divisions Zygomycota, Deuteromycota and Ascomycota. Some moulds cause disease or food spoilage; others play an important role in biodegradation or in the production of various foods, beverages, antibiotics and enzymes.
There are hundreds of known species of moulds that include opportunistic pathogens, saprophytes and thermophiles. Like all other fungi, moulds derive energy from the organic matter on which they grow. Typically, moulds secrete hydrolytic enzymes, mainly from the hyphal tips. These enzymes degrade complex biopolymers (i.e. starch, cellulose and lignin) into simpler substances that can be absorbed by the hyphae. In this way moulds play a major role in causing decomposition of organic material, enabling the recycling of nutrients throughout ecosystems. Many moulds also secrete mycotoxins that, together with hydrolytic enzymes, inhibit the growth of competing microbes.
Moulds reproduce through small spores that may contain a single nucleus or be multinucleate. Mould spores can be asexual or sexual; many species can produce both types. Mould spores may remain airborne indefinitely, may attach to clothing or fur and survive extreme of temperature and pressure. Although moulds grow on dead organic matter everywhere in nature, their presence is visible to the unaided eye only when mould colonies grow. A mould colony does not consist of discrete microbes but of an interconnected network of hyphae called a mycelium. Nutrients may be transported throughout the mycelium.
Few moulds can grow at 4°C, the temperature within a typical household refrigerator, or less. When conditions do not enable growth, moulds may remain alive in a dormant state, within a large range of temperatures. The different mould species vary enormously in their tolerance to temperature and humidity. Certain moulds can survive harsh conditions for example refrigeration, highly acidic solvents, anti-bacterial soap and even petroleum products. Xerophilic moulds use the humidity in the air as their only water source; other moulds need more moisture.
10.3 Penicillium
Penicillium is an ascomycetous fungus and is of major importance in natural environment as well as food and drug production. Members of this genus produce penicillin, an antibiotic that kills or stops the growth of certain bacteria. According to the Dictionary of the Fungi, it is the widespread genus that contains over 300 species. Penicillium has aseptate vegetative mycelia that penetrate the substrate and then produce aerial hyphae on which conidiophores develop. These condiophores may be branched and have brush like heads bearing spores clusters and each is form a chain of conidia. With the production of conidia, colonies become green, grey green, blue green and yellow green.
Penicillium is classified as a genus of anamorphic fungi in the division Ascomycota (order Eurotiales, class Eurotiomycetes, family Trichocomaceae). The name is derived from the Latin root Penicillium, meaning ‘painter's brush’, and refers to the chains of conidia that resemble a broom.
The thallus (mycelium) typically
consists of a highly branched network of multinucleate, septate,
usually colourless hyphae. Many-branched conidiophores
sprout on the mycelia, bearing individually constricted conidiospores.
The conidiospores are the main dispersal route of
fungi, and often are green in colour. Sexual reproduction involves the
production of ascospores, commencing with the fusion
of an archegonium and an antheridium,
with sharing of nuclei. The irregularly distributed asci
contain eight unicellular ascospores each.
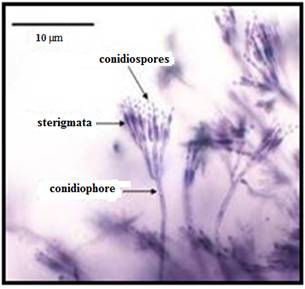
Fig. 10.1 Pencillium spp
Penicillium roqueforti is a common saprotrophic fungus of family Trichocomaceae. It is widespread in nature and can be isolated from soil and decaying organic matter. The major industrial use of this fungus is the production of blue cheeses, flavoring agents, antifungals, polysaccharides, proteases and other enzymes. The fungus has been a constituent of Roquefort and other blue veined cheeses.
Asymmetric group i.e. more than one branch in a conidiophore and the branching is asymmetrical. Produces blue-green spreading colonies on malt agar gradually change to a darker green color. Colonies are smooth velvety appearance with irregular margins of radiating lines of conidiophores. Conidiophores are rough, globose and smooth. The toxins produced are Roquefortin, mycophenolic acid, PR toxin,
Penicillium camemberti is a species of fungus used in the production of Camembert and Brie cheeses, on which colonies of P. camemberti form a hard, white crust. It is responsible for giving these cheeses their distinctive taste.
10.4 Geotrichum
Geotrichum candidum is a fungus or mould that can act as a plant pathogen causing sour rot on peach, nectarine, tomato and carrot. It is also widely used in the production of many dairy products including many natural rind cheeses such as Camembert and other bloomy rind cheese. It is also found in the Nordic dairy product villi and is responsible for the velvety surface of the product. It forms white colonies, yeast like form true mycelium, breaks to form arthrospore that are cylindrical with rounded ends.
10.5 Aspergillus
All Aspergillus species are highly aerobic and are found in oxygen-rich environments, where these commonly grow as moulds on the surface of a substrate, as a result of high oxygen tension. Commonly, fungi grow on carbon-rich substrates like monosaccaharides (i.e. glucose) and polysaccaharides (i.e. amylose). Aspergillus is common contaminants of starchy foods.
In addition to growth on carbon sources, many species of Aspergillus demonstrate oligotrophy, where these grow in nutrient-depleted environments, or environments where there is a complete lack of key nutrients. A. niger is a prime example of this; it can be found growing on damp walls, as a major component of mildew.
Aspergillus niger is one of the most common species of Aspergillus. It causes a disease called black mould on certain fruits and vegetables such as grapes, onions, and peanuts, and is a common contaminant of food. It is ubiquitous in soil and is commonly reported from indoor environments, where its black colonies can be confused with those of Stachybotrys (black mould).
Aspergillus flavus is a common mould in the environment that causes problems in stored grains. It can also be a human pathogen, associated with aspergillosis of the lungs and sometimes causing corneal, otomycotic, and nasoorbital infections. Many strains produce significant quantities of aflatoxin, a carcinogenic and acutely toxic compound. A. flavus spores are allergenic.
A. flavus grows as a yellow-green mould in culture. Like other Aspergillus species, it produces a distinctive conidiophores composed of a long stalk supporting an inflated vesicle. Conidiogenous cells on the vesicle produce the conidia. Many strains of A. flavus exhibit a greenish fluorescence under UV light that is correlated with levels of aflatoxin production.
10.6 Yeasts
Yeasts are eukaryotes classified in the kingdom Fungi, with 1,500 species estimated to be 1% of total fungal species. Most reproduce asexually, and many do so by an asymmetric division called budding. Yeasts are unicellular, although some species with yeast forms may become multicellular through the formation of a string of connected budding cells known as pseudohyphae, or false hyphae. Yeasts can vary greatly in size depending on the species, typically measuring 3–4 µm in diameter, although some yeast can reach over 40 µm.
Saccharomyces cerevisiae converts carbohydrates to carbon dioxide and alcohols – for years carbon dioxide has been used in baking and alcohol in beverages. Other species of yeast, like Candida albicans, are opportunistic pathogens and can cause infections in humans. Yeasts have been used to generate electricity in microbial fuel cells, and produce ethanol for the biofuel industry these days.
Yeasts do not form a single taxonomic or phylogenetic grouping. The term ‘yeast’ is often taken as a synonym for Saccharomyces cerevisiae, but the phylogenetic diversity of yeasts is shown by their placement in two separate phyla: the Ascomycota and the Basidiomycota. The budding yeasts (‘true yeasts’) are classified in the order Saccharomycetales.
Yeasts are unicellular-spherical/ ovoid, pseudomycelium, vegetative reproduction by budding, ascospores–1 to 4 per ascus.
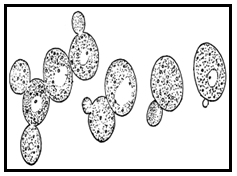
Fig. 10.2 Budding yeasts
Yeasts multiply asexually by budding, where a bud has formed on a cell. As many as to bud scars are found on a single cell. During budding nucleus divides by construction and a portion of it enters the bud along with other organelles. The cytoplasmic connection is formed by laying down of cell wall material.
Under appropriate conditions–the yeasts forms asci. The cytoplasm of the cell differentiates into four thickened wall spherical spores, although the number of spores can be less. The cells from which asci develop are diploid and nuclear divisions that precede spore formation are meiotic.
10.7 Saccharomyces
Saccharomyces cerevisiae is the most useful yeast, having been used in baking and brewing. It was originally isolated from and is one of the most studied eukaryotic micro-organisms like Escherichia coli as a prokaryote. S. cerevisiae cells are round to ovoid, 5–10 µm in diameter and reproduces by budding.
It is known as Baker’s yeast, isolated from kefir. Cells - globose, subglobose, ellipsoidal/ cylindrical singles, pairs, short chains/ clusters. Ascospores - globose to short ellipsoidal, 1-4 per ascus, do not liberate, do not ferment lactose and nitrate is not assimilated.
10.8 Kluyveromyces
10.8.1 Kluyveromyces fragilis
· Used in Kefir and Koumiss fermentation.
· Cells–subglobose, ellipsoidal to cylindrical single or pairs, ascospores–1 to 4 per ascus cresentiform or reniform.
· Lactose may be fermented by some of the strains while nitrate is not assimilated.
10.8.2 Kluyveromyces lactis
· Associated with yoghurt, isolated from milk, gassy cheese, Italian cheese, and cream and butter milk.
· Cells–spherical, ellipsoidal or occasionally cylindrical clusters, singles, pairs and occasionally clusters
· Ascospores–1 to 4, spherical to ellipsoidal, readily released, lactose fermented but nitrate is not assimilated
10.9 Candida
10.9.1 Candida kefir
It is associated with kefir, buttermilk and cheese.
· Morphologically variable, budding unicellular to pseudomycelium or true mycelium
· Cells–Ovoid short/ long, Reproduction – budding or fission, Colonies–off white to cream, lactose may be fermented but nitrate is not assimilated.
10.9.2 Candida lacticondensi – (Torulopsis lactiscondensi)
Cells are ovoid, budding, the typical colonies on selective media are–off-white, cream and yellowish or brownish, mycelium not formed or rudimentary mycelium is rarely found. Lactose is not utilized as carbon source nor do they assimilate nitrate.
10.10 Bacteriophage
These are the viruses that infect bacteria. These are the quintessential parasites of the living kingdoms. ‘Phage’ literally means eating just like phagocyte (to swallow). These are widely distributed in nature and most abundant in intestinal contents of animals and were first invented by Twort in 1915. Typical bacteriophage has a structure consisted of:
· Head–enclosing nucleic acids in protein sheath.
· Tail–a hollow tube of proteins bearing tail plate and fibers.
All viruses contain nucleic acid, either DNA or RNA (but not both together), and a protein coat, that encases the nucleic acid. Some viruses are also enclosed by an envelope of fat and protein molecules. Without a host cell, viruses cannot carry out the life-sustaining functions or reproduce (Fig. 10.3).
These cannot synthesize proteins, because these lack ribosomes and must use the ribosomes of host cells to translate viral messenger RNA into viral proteins. Viruses cannot generate or store energy in the form of adenosine triphosphate, but have to derive it and all other metabolic functions, from the host cell. These also parasitize the cell for basic building materials, (i.e. amino acids, nucleotides, and lipids). These are significant from dairy or food processing point of view as; phages can attack lactic acid bacteria and lyses them by multiplying inside the host leading to the release of many phages that can re-infect the new cells in culture. This results in the failure of starters to act and bring about changes during preparation of fermented products.
· Bacteriophages are highly host specific and the rotation of starter cultures may help to control the problem to some extent.
· Ca++ ion deficient medium for maintaining starter cultures prevent phage attack as the ion helps in phage adsorption to host cell.
· Genetic manipulations to construct phage resistant strains.
Typical Lactococcus lactis ssp lactis phage might have 40-90nm head and 100-120nm tail. Bacteriophages can persist for a long time in dried up whey and on utensils.
Phages attach themselves to living cells and as these divide phages also multiply. The infective material is injected (nucleic acid) through hollow tail of the phage particle into bacterial cell. The rate of phage proliferation is greater than that of bacterium. Therefore, a point is reached at which lysis of host cell begins. Lysis is preceded by swelling of the bacteria to 6-16 times than their normal size. Phage action may cause a number of variations in the properties of bacteria. Irreversible adsorption of phage to the cell triggers a number of steps that lead to the transport of viral nucleic acids into cytoplasm and nucleic acid gets metabolically active transforming cellular function to one of synthesizing viral components. As the phage assembly completed the lysis of host cell occur. All this occur nearly in one generation time of the host.